Den visuelle cortex (V1): Struktur, funktion og banebrydende forskning
Den visuelle cortex (V1): Dyk ned i struktur, funktion og banebrydende forskning — fra Nobel-opdagelser til moderne neuroteknikker og synets grundlæggende byggesten.
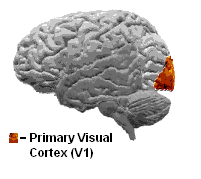
Den visuelle cortex er en del af hjernen, som gør det muligt at se. Den er relativt tynd - mellem 1,5 og 2 mm hos mennesker. Hos aber og aber fylder den visuelle cortex en stor del af deres hjerne. Fysisk set ligger den visuelle cortex bagerst i hjernen i baghovedlappen.
David Hubel og Torsten Wiesel forskede i mange år i den visuelle cortex. De vandt Nobelprisen i fysiologi eller medicin i 1981 for deres opdagelser om informationsbehandling i det visuelle system.
- I 1960'erne og 1970'erne arbejdede de med at undersøge, hvordan det visuelle system udviklede sig. De arbejdede med de dele af hjernens visuelle cortex, som modtager signaler fra højre eller venstre øje.
- I deres arbejde beskrives det, hvordan signaler fra øjet behandles af hjernen til at generere kantdetektorer, bevægelsesdetektorer, stereoskopiske dybdedetektorer og farvedetektorer. Disse er byggestenene i den visuelle scene.
Forskning i den primære visuelle cortex kan involvere registrering af aktionspotentialer fra elektroder i hjernen hos katte, fritter, rotter, mus eller aber. Alternativt kan signalerne registreres uden for dyret ved hjælp af EEG, MEG eller fMRI. Disse teknikker indsamler oplysninger uden at trænge ind i hjernen.
Billedgalleri
8 Billeder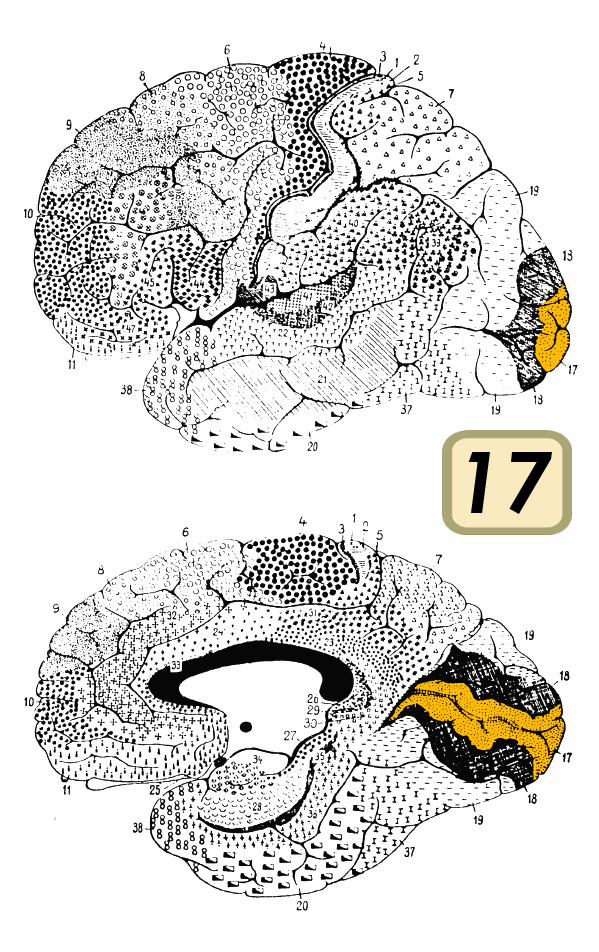






Struktur og lagopdeling
Den primære visuelle cortex kaldes ofte V1 og er organiseret i seks tydelige lag (1–6). Hver lagtype har forskellige celletyper og forbindelser:
- Layer 4 (særligt 4C) modtager hovedparten af input fra lateral geniculate nucleus (LGN) i thalamus.
- Superficielle lag (2/3) sender information videre til næste visuelle områder (f.eks. V2, V4, MT/V5) og indeholder cytochrome-oxidase-blobs for farvebehandling.
- Dybere lag (5/6) projicerer tilbage til thalamus og til andre hjernestrukturer.
Funktionel organisation: receptive fields og celletype
Hver nervecelle i V1 har et receptivt felt — et område i synsfeltet, hvor stimuli påvirker cellens aktivitet. De centrale funktionsmæssige egenskaber omfatter:
- Simple celler: reagerer på kanter eller streger med bestemt orientering og position inden for receptive field.
- Complex celler: reagerer på orienterede bevægelser uafhængigt af præcis position.
- End-stopped celler: hæmmet af langstrakte stimuli og sensitive for ender eller vinkler.
Rumlig organisation: retinotopi og cortical magnification
V1 er retinotopisk organiseret: naboområder i nethinden kortlægges på naboområder i cortex. Fovea (det centrale synsfelt) har en uforholdsmæssigt stor repræsentation i V1—et fænomen kaldet cortical magnification. Hos mennesker ligger meget af V1 i calcarine fissur (en dyb fure), hvoraf foldningen gør kortlægningen kompleks, men funktionelt præcis.
Udvikling og plasticitet
Hubel og Wiesel viste, at normal visuel stimulation i en kritisk periode tidligt i livet er nødvendig for korrekt udvikling af ocular dominance-kolonner og binokulær funktion. Hvis et øje dækkes i denne periode (f.eks. ved øjenlidelse hos unge dyr), kan cortex reorganisere sig, hvilket fører til amblyopi (»dovent øje«). Denne plastiske evne aftager med alderen, men voksenplasticitet og genoptræning kan i visse tilfælde forbedre funktion.
Metoder i moderne forskning
Teknikker, der anvendes i V1-forskning, spænder fra invasive til ikke-invasive:
- Single-unit og multi-unit elektrofysiologi (elektroder) for at måle aktionspotentialer i dyr (som katte, rotter, mus eller aber).
- In vivo two-photon calcium imaging for at registrere aktivitet i store cellepopulationer med god rumlig opløsning.
- Optogenetik for at stimulere eller hæmme specifikke celletyper med lys.
- Non-invasive metoder som EEG, MEG og fMRI til at studere menneskers visuelle funktion og kortlægge aktivitet uden kirurgi.
- Computational modelling og kunstige neurale netværk (f.eks. convolutional neural networks) inspirerer af V1-mekanismer (Gabor-lignende filtre) og genererer hypoteser om hierarkisk visuel behandling.
Artsforskelle
Den visuelle cortex varierer mellem arter. Hos primater (inkl. mennesker) er V1 stor og komplekst foldet; hos gnavere er V1 ofte fladere og mindre udbygget, men grundlæggende funktioner (orienterings- og bevægelsessensitivitet) kan stadig findes. Valget af forsøgsdyr afhænger af hvilke spørgsmål, der efterspørges — f.eks. detaljeret kolonnearkitektur i aber vs. genetiske værktøjer i mus.
Klinisk betydning og anvendelser
Skader i V1 (f.eks. ved slagtilfælde eller traume) kan føre til synsfeltstab eller cortical blindhed. Forståelse af V1 er central for behandling og teknologiske løsninger:
- Visuelle proteser (retinale eller cortikale implants) forsøger at genskabe elementær visuel information ved at stimulere neuroner.
- Diagnose og behandling af amblyopi og andre udviklingsrelaterede synsforstyrrelser bygger på kendskab til kritiske perioder i V1.
- Forskning i V1 har inspireret billedbehandling og computer vision, hvilket forbedrer algoritmer til kant- og mønstergenkendelse.
Banebrydende og aktuel forskning
Ud over Hubel og Wiesel går moderne forskning videre med:
- Kartlægning af mikroarkitektur (orientation pinwheels, kolonner) med høj opløsning.
- Studier af hvordan naturlige bevægelser og komplekse scener kodes i populationsaktivitet.
- Integration af optogenetik, calcium-imaging og storskala elektrofysiologi for at forstå netværksdynamik.
- Udvikling af neurale proteser og BMI (brain–machine interfaces) med mål om at genoprette syn eller tilbyde nye sensoriske inputs.
Den primære visuelle cortex er dermed både et modelområde for forståelse af sensorisk kodning og et centralt felt for kliniske og teknologiske anvendelser. Forskning fortsætter med at kortlægge, hvordan simple filtre i V1 kombineres til komplekse og meningsfulde visuelle oplevelser.

Primær visuel cortex
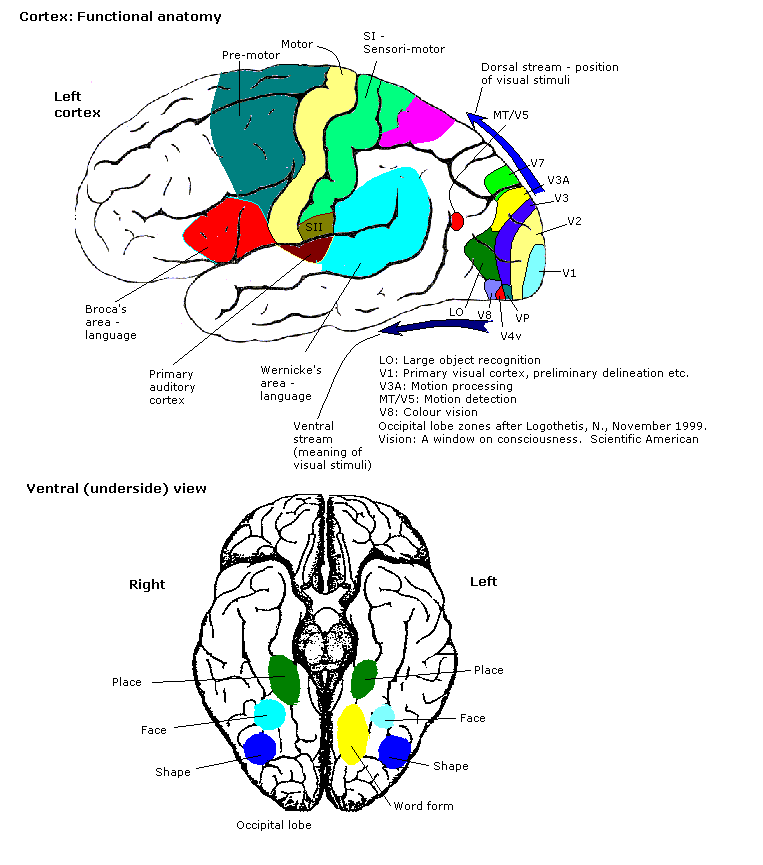
Den primære visuelle cortex (V1) er det bedst undersøgte visuelle område i hjernen. Det er her, at beskederne kommer fra de laterale geniculære kerner, som er relæstationer for information fra nethinden. Hver lateral geniculær kerne får signaler fra det modsatte synsfelt.
Hver V1 sender information til to primære baner, kaldet den ventrale strøm og den dorsale strøm.
- Den ventrale strøm begynder ved V1, går gennem det visuelle område V2, derefter gennem det visuelle område V4 og til den nedre temporale cortex (IP cortex). Den ventrale strøm, der undertiden kaldes "What Pathway", er forbundet med formgenkendelse og objektrepræsentation. Den er også forbundet med lagring af langtidshukommelse.
- Den dorsale strøm begynder ved V1, går gennem det visuelle område V2, derefter til det dorsomediale område (DM/V6) og det visuelle område MT (Middle temporal/V5) og til den bageste parietale cortex. Den dorsale strøm, der undertiden kaldes "Where Pathway" eller "How Pathway", er forbundet med bevægelse, repræsentation af objektplaceringer og kontrol af øjne og arme, især når visuelle oplysninger bruges til at styre øjenbevægelser eller række ud.
Spørgsmål og svar
Spørgsmål: Hvad er den visuelle cortex?
A: Den visuelle cortex er en del af hjernen, der gør det muligt at se. Den er placeret i occipitallappen bagest i hjernen og er relativt tynd, mellem 1,5 og 2 mm hos mennesker.
Spørgsmål: Hvem har forsket i den visuelle cortex?
Svar: David Hubel og Torsten Wiesel forskede i mange år i den visuelle cortex. De vandt i 1981 Nobelprisen i fysiologi eller medicin for deres opdagelser om informationsbehandling i det visuelle system.
Spørgsmål: Hvilken slags forskning foretog de?
A: I 1960'erne og 1970'erne arbejdede de med at undersøge, hvordan det visuelle system udviklede sig. De arbejdede med dele af hjernens visuelle cortex, som modtager signaler fra begge øjne, og beskrev, hvordan signaler fra disse øjne behandles af hjernen for at generere kantdetektorer, bevægelsesdetektorer, stereoskopiske dybdedetektorer og farvedetektorer - byggestenene i en visuel scene.
Spørgsmål: Hvordan kan forskerne undersøge aktiviteten i den primære visuelle cortex?
A: Forskning i primær visuel cortex-aktivitet kan bestå i at optage aktionspotentialer fra elektroder i et dyrs hjerne (katte, fritter, rotter, mus eller aber). Alternativt kan signalerne registreres uden for dyret ved hjælp af EEG-, MEG- eller fMRI-teknikker, som indsamler oplysninger uden at trænge ind i dyrets hjerne.
Spørgsmål: Hvor tyk er menneskets visuelle cortex?
Svar: Menneskets visuelle cortex er relativt tynd - mellem 1,5 og 2 mm tyk.
Spørgsmål: Hvilken pris vandt Hubel og Wiesel for deres opdagelser om informationsbehandling i det visuelle system?
Svar: David Hubel og Torsten Wiesel vandt i 1981 Nobelprisen i fysiologi eller medicin for deres opdagelser om informationsbehandling i det visuelle system.
Relaterede artikler
Forfatter
AlegsaOnline.com Den visuelle cortex (V1): Struktur, funktion og banebrydende forskning Leandro Alegsa
URL: https://da.alegsaonline.com/art/105637
Kilder
- sciencedirect.com : sciencedirect.com/science/article/pii/S0042698910003743